Archives
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2018-07
-
Typically upon chronic agonist exposure GPCRs undergo desens
2021-11-10
Typically, upon chronic agonist exposure GPCRs undergo desensitization and internalization resulting in a loss of receptor responsiveness over time (Drake et al., 2006, Kelly et al., 2008). However, not all GPCR systems conform to this model of acute agonist-mediated regulation. Some receptors have
-
Our overall goal is to develop selective compounds
2021-11-10
Our overall goal is to develop selective compounds for low-affinity/high-capacity transporters inhibitors (i.e. OCT1–3 and PMAT), and provide the field with much needed, specific pharmacological tools to study these transporters. In our initial efforts, we synthesized halogen substituted analogs and
-
br Experimental section br Introduction Fasting hyperglycaem
2021-11-10
Experimental section Introduction Fasting hyperglycaemia in all forms of diabetes mellitus occurs primarily as a result of upsurge in hepatic glucose synthesis (HGS) [1], [2]. Two specific enzymes, glucokinase (GK) and glucose-6 phosphatase (Glu-6-Pase) play crucial role in hepatic glucose pro
-
Genomic studies have afforded new insight into
2021-11-10
Genomic studies have afforded new insight into the influence of rhein structure on GR binding. An important clue came from genome-wide mapping of chromatin structure using DNase I hypersensitive site sequencing (DNase-seq), and comparing the locations of open chromatin to GBSs [10]. DNase-seq in th
-
By association a possible mechanism which
2021-11-10
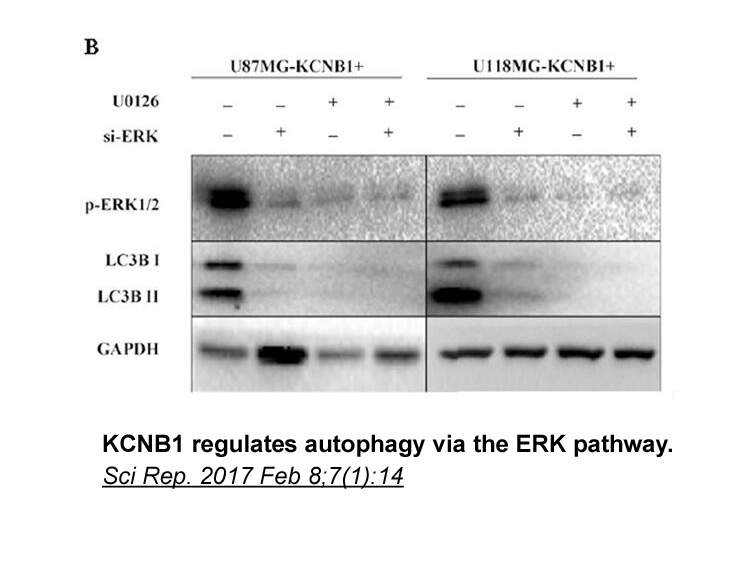
By association, a possible mechanism which may underlie the progression of seizures is long-term potentiation (LTP). Indeed, LTP shares many common mechanisms with seizures activity both anatomically and physiologically, and has been implicated as a mechanism of acute epileptogenesis (Wasterlain et
-
br Role of FPRs in regulating
2021-11-10
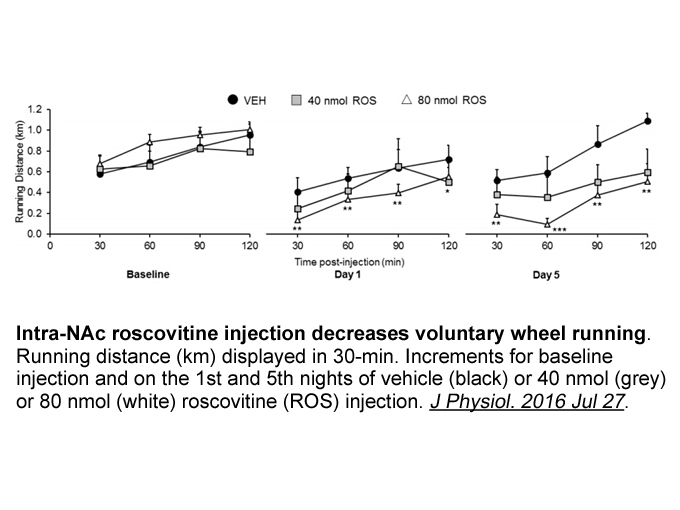
Role of FPRs in regulating the inflammatory response Annexin I is a glucocorticoid-regulated protein possessing both pro- and anti-inflammatory activity that might be mediated, in part, by FPR activation [35]. Expressed in a variety of cell types, annexin I is particularly abundant in neutrophils
-
A frequent complication of SIRS
2021-11-10
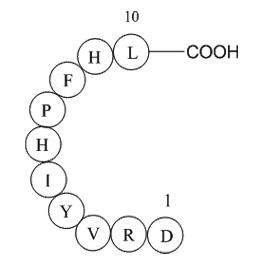
A frequent complication of SIRS and sepsis is the development of increased airway resistance and lung inflammation [2], [3]. For example, Uhlig et al. [3] demonstrated that rat lungs exposed to endotoxin present increased airway resistance due to constriction of terminal adenosine receptor antagonis
-
The FAAH inhibitor activity of
2021-11-10
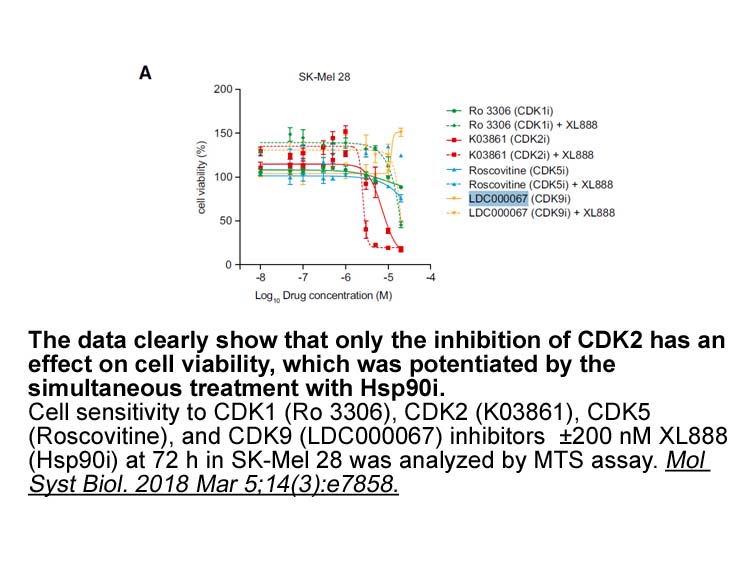
The FAAH inhibitor activity of the 3-(5-(ethoxycarbonyl)-1-phenyl-1H-pyrazol-3-yl)phenyl cyclohexylcarbamates was investigated by preparation of compounds (43–52, Table 3). These new compounds displayed an important loss of activity in comparison with the corresponding regioisomers 5-phenyl compound
-
br Cellular transportation of nanoparticles
2021-11-10
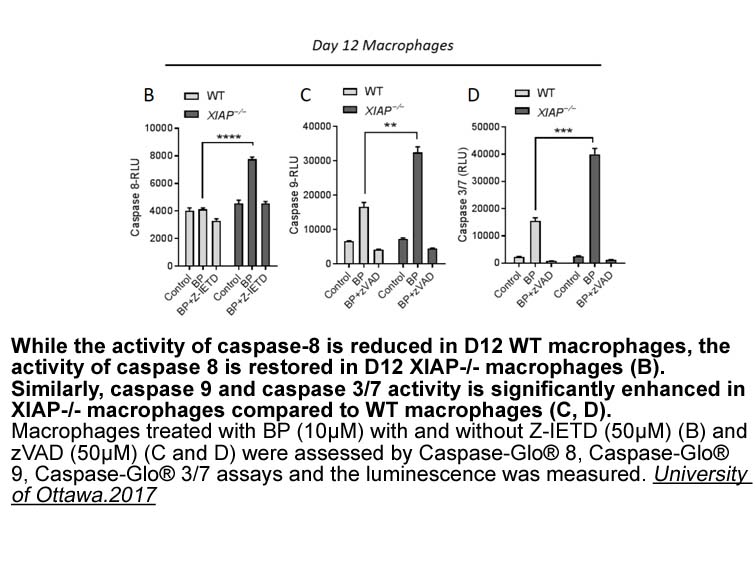
Cellular transportation of nanoparticles and nanocomplexes Endocytosis and exocytosis are critically important phenomena in cellular metabolism, which involve the complex vesicular system. They deal with major cellular transportation activities including membrane homeostasis by transporting prote
-
Our extensive preliminary data suggest that the loss of p
2021-11-10
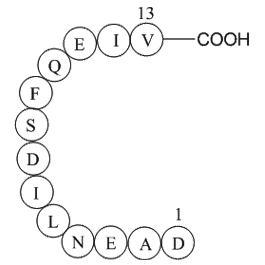
Our extensive preliminary data suggest that the loss of p300 (and resulting H3K27ac) is related to inorganic arsenic related diseases. Therefore, it may open a new avenue for alleviating the consequence of H3K27ac, for example, via boosting CBP enzymatic activity for at least partially compensating
-
br Hepatitis C virus HCV infection is a major cause
2021-11-09
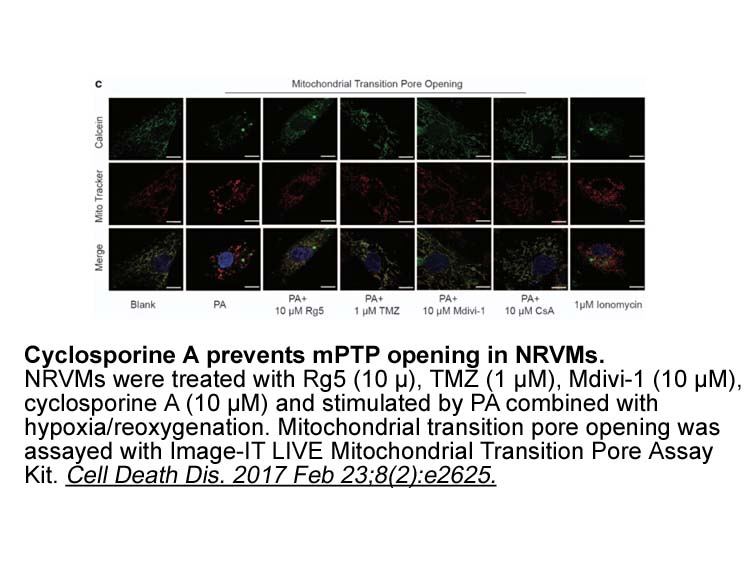
Hepatitis C virus (HCV) infection is a major cause of chronic liver disease that can lead to cirrhosis and hepatocellular carcinoma. It is estimated that nearly 200 million individuals worldwide are currently infected with HCV and it is the leading cause of liver transplants. The current standard
-
Finally we investigated increasing basicity as a
2021-11-09
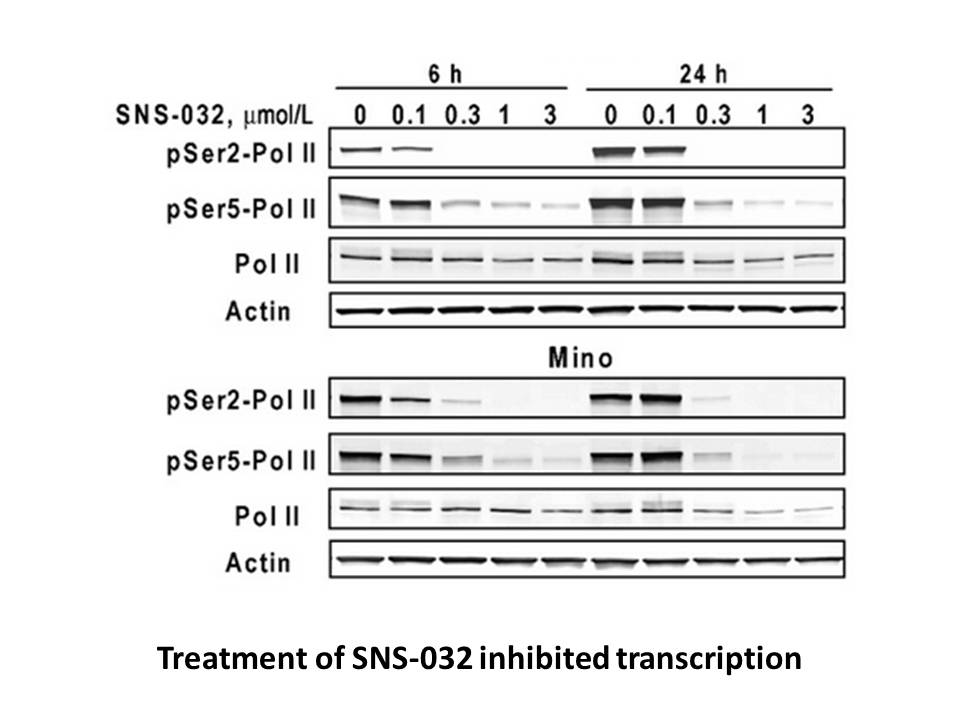
Finally, we investigated increasing basicity as a way to improve solubility. We expected that conversion of the ether linkage of to an -methyl group could improve solubility. More than expected, fccp demonstrated significant improvement of values of solubility for the two media, especially for ac
-
The differentially expressed proteins during
2021-11-09
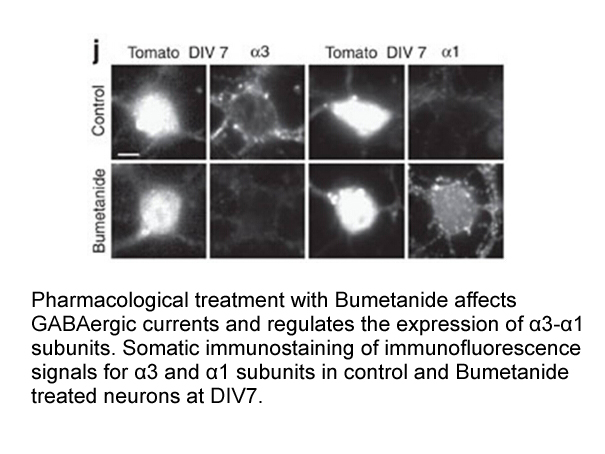
The differentially expressed proteins during somatic embryogenesis are described as the proteins that are induced by oxidative stress and respond to higher cell division activity including ascorbate peroxidase, dehydroascorbate reductase, glutathione transferase and mitochondrial manganese superoxid
-
One possible pathway summarizing the overall action involves
2021-11-09
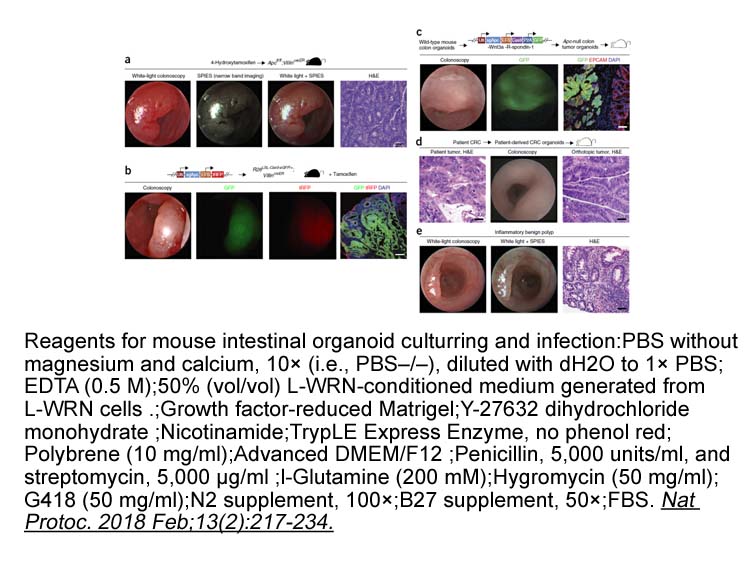
One possible pathway summarizing the overall action involves NMDA receptor activation – Ca2+ influx – protein kinase A (PKA) activation – ERK phosphorylation and subsequent synaptic plasticity (Thomas and Huganir, 2004). A few studies have also shown modulation of synaptic glutamate transporters in
-
dl 1559 By regulation of synaptic glutamate concentration
2021-11-09
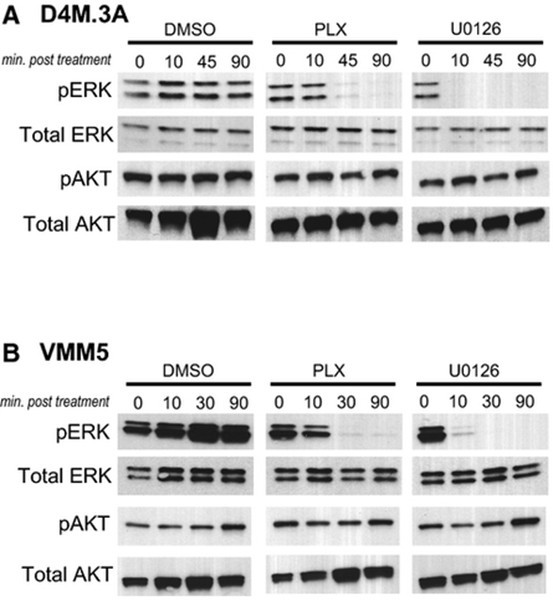
By regulation of synaptic glutamate concentration, glutamate transporters play an important role in limiting glutamate signaling and controlling the activation of glutamate receptors (Rimmele and Rosenberg, 2016). Reduced glutamate transport is thought to underlie the pathogenesis of numerous neurol
15307 records 553/1021 page Previous Next First page 上5页 551552553554555 下5页 Last page